

Does An Organism Have A Way Of Repairing Dna
Deoxyribonucleic acid integrity is always under attack from environmental agents like skin cancer-causing UV rays. How do DNA repair mechanisms detect and repair damaged Dna, and what happens when they fail?

Considering DNA is the repository of genetic information in each living cell, its integrity and stability are essential to life. Deoxyribonucleic acid, however, is not inert; rather, information technology is a chemical entity subject area to assault from the environs, and any resulting harm, if not repaired, will lead to mutation and possibly disease. Possibly the best-known example of the link between ecology-induced DNA harm and disease is that of skin cancer, which tin be acquired by excessive exposure to UV radiation in the form of sunlight (and, to a lesser degree, tanning beds). Another example is the harm caused by tobacco fume, which tin can lead to mutations in lung cells and subsequent cancer of the lung. Beyond environmental agents, Dna is as well subject to oxidative damage from byproducts of metabolism, such every bit free radicals. In fact, information technology has been estimated that an individual cell tin suffer up to 1 million DNA changes per solar day (Lodish et al., 2005).
In addition to genetic insults caused past the environment, the very process of DNA replication during cell partition is prone to mistake. The rate at which Deoxyribonucleic acid polymerase adds incorrect nucleotides during Deoxyribonucleic acid replication is a major factor in determining the spontaneous mutation rate in an organism. While a "proofreading" enzyme normally recognizes and corrects many of these errors, some mutations survive this process. Estimates of the frequency at which man Deoxyribonucleic acid undergoes lasting, uncorrected errors range from 1 ten 10-4 to 1 x ten-6 mutations per gamete for a given gene. A charge per unit of 1 x 10-6 ways that a scientist would expect to find ane mutation at a specific locus per i million gametes. Mutation rates in other organisms are frequently much lower (Table 1).
One way scientists are able to estimate mutation rates is by considering the rate of new dominant mutations found at different loci. For example, by examining the number of individuals in a given population who were diagnosed with neurofibromatosis (NF1, a disease acquired by a spontaneous—or noninherited—dominant mutation), scientists determined that the spontaneous mutation rate of the gene responsible for this illness averaged 1 10 10-4 mutations per gamete (Crowe et al., 1956). Other researchers have establish that the mutation rates of other genes, like that for Huntington'due south disease, are significantly lower than the rate for NF1. The fact that investigators take reported different mutation rates for different genes suggests that certain loci are more prone to harm or fault than others.
DNA Repair Mechanisms and Man Disease

DNA repair processes exist in both prokaryotic and eukaryotic organisms, and many of the proteins involved have been highly conserved throughout development. In fact, cells accept evolved a number of mechanisms to notice and repair the various types of impairment that can occur to DNA, no matter whether this harm is caused past the environment or by errors in replication. Because Dna is a molecule that plays an agile and critical role in cell segmentation, command of DNA repair is closely tied to regulation of the prison cell cycle. (Call back that cells transit through a cycle involving the 10001, S, Yard2, and 1000 phases, with DNA replication occurring in the South stage and mitosis in the M stage.) During the cell cycle, checkpoint mechanisms ensure that a cell's Dna is intact before permitting Deoxyribonucleic acid replication and cell division to occur. Failures in these checkpoints can pb to an accumulation of damage, which in turn leads to mutations.
Defects in DNA repair underlie a number of human genetic diseases that touch a wide diversity of torso systems just share a constellation of mutual traits, most notably a predisposition to cancer (Tabular array ii). These disorders include ataxia-telangiectasia (AT), a degenerative motor condition acquired by failure to repair oxidative damage in the cerebellum, and xeroderma pigmentosum (XP), a condition characterized by sensitivity to sunlight and linked to a defect in an of import ultraviolet (UV) damage repair pathway. In improver, a number of genes that take been implicated in cancer, such as the RAD group, have also been determined to encode proteins critical for Dna damage repair.
UV Damage, Nucleotide Excision Repair, and Photoreactivation

Equally previously mentioned, one important DNA damage response (DDR) is triggered by exposure to UV calorie-free. Of the iii categories of solar UV radiation, only UV-A and UV-B are able to penetrate World's atmosphere. Thus, these 2 types of UV radiation are of greatest concern to humans, particularly as continuing depletion of the ozone layer causes college levels of this radiation to reach the planet's surface.
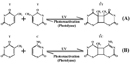
UV radiation causes two classes of DNA lesions: cyclobutane pyrimidine dimers (CPDs, Figure 1) and vi-4 photoproducts (6-iv PPs, Figure 2). Both of these lesions distort DNA's structure, introducing bends or kinks and thereby impeding transcription and replication. Relatively flexible areas of the DNA double helix are most susceptible to damage. In fact, 1 "hot spot" for UV-induced harm is found inside a commonly mutated oncogene, the p53 factor.
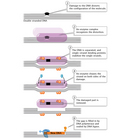
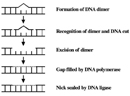
CPDs and half dozen-4 PPs are both repaired through a process known as nucleotide excision repair (NER). In eukaryotes, this complex process relies on the products of approximately thirty genes. Defects in some of these genes have been shown to cause the human disease XP, every bit well every bit other conditions that share a risk of skin cancer that is elevated about a thousandfold over normal. More specifically, eukaryotic NER is carried out by at least 18 protein complexes via four discrete steps (Figure three): detection of damage; excision of the section of Dna that includes and surrounds the error; filling in of the resulting gap by DNA polymerase; and sealing of the nick between the newly synthesized and older DNA (Figure four). In bacteria (which are prokaryotes), still, the process of NER is completed by only three proteins, named UvrA, UvrB, and UvrC.
Leaner and several other organisms also possess some other mechanism to repair UV impairment called photoreactivation. This method is oft referred to as "lite repair," because it is dependent on the presence of light energy. (In comparison, NER and well-nigh other repair mechanisms are oft referred to as "nighttime repair," every bit they do not require lite as an energy source.) During photoreactivation, an enzyme called photolyase binds pyrimidine dimer lesions; in addition, a 2nd molecule known as chromophore converts light energy into the chemical energy required to directly revert the afflicted area of DNA to its undamaged form. Photolyases are found in numerous organisms, including fungi, plants, invertebrates such as fruit flies, and vertebrates including frogs. They do not appear to exist in humans, however (Sinha & Hader, 2002).
Additional DNA Repair mechanisms
NER and photoreactivation are not the only methods of DNA repair. For instance, base excision repair (BER) is the predominant mechanism that handles the spontaneous DNA damage caused by free radicals and other reactive species generated by metabolism. Bases tin go oxidized, alkylated, or hydrolyzed through interactions with these agents. For case, methyl (CH3) chemical groups are frequently added to guanine to grade 7-methylguanine; alternatively, purine groups may exist lost. All such changes result in abnormal bases that must exist removed and replaced. Thus, enzymes known as Deoxyribonucleic acid glycosylases remove damaged bases by literally cutting them out of the DNA strand through cleavage of the covalent bonds between the bases and the sugar-phosphate courage. The resulting gap is then filled by a specialized repair polymerase and sealed by ligase. Many such enzymes are institute in cells, and each is specific to certain types of base alterations.
Still another form of Deoxyribonucleic acid harm is double-strand breaks, which are caused by ionizing radiation, including gamma rays and 10-rays. These breaks are highly deleterious. In addition to interfering with transcription or replication, they tin atomic number 82 to chromosomal rearrangements, in which pieces of 1 chromosome become attached to another chromosome. Genes are disrupted in this process, leading to hybrid proteins or inappropriate activation of genes. A number of cancers are associated with such rearrangements. Double-strand breaks are repaired through one of two mechanisms: nonhomologous end joining (NHEJ) or homologous recombination repair (HRR). In NHEJ, an enzyme called DNA ligase IV uses overhanging pieces of Deoxyribonucleic acid adjacent to the suspension to join and fill in the ends. Boosted errors can be introduced during this process, which is the example if a cell has not completely replicated its DNA in training for division. In contrast, during HRR, the homologous chromosome itself is used as a template for repair.
Mutations in an organism's Deoxyribonucleic acid are a part of life. Our genetic code is exposed to a variety of insults that threaten its integrity. Merely, a rigorous system of checks and balances is in place through the Deoxyribonucleic acid repair machinery. The errors that slip through the cracks may sometimes be associated with affliction, but they are also a source of variation that is acted upon by longer-term processes, such as evolution and natural option.
References and Recommended Reading
Branze, D., & Foiani, M. Regulation of Deoxyribonucleic acid repair throughout the jail cell cycle. Nature Reviews Molecular Cell Biology 9, 297–308 (2008) doi:10.1038/nrm2351.pdf (link to article)
Crowe, F. W., et al. A Clinical, Pathological, and Genetic Report of Multiple Neurofibromatosis (Springfield, Illinois, Charles C. Thomas, 1956)
Lodish, H., et al. Molecular Biology of the Jail cell, 5th ed. (New York, Freeman, 2004)
Sinha, R. P., & Häder, D. P. UV-induced DNA damage and repair: A review. Photochemical and Photobiological Sciences 1, 225–236 (2002)

Does An Organism Have A Way Of Repairing Dna,
Source: https://www.nature.com/scitable/topicpage/dna-damage-repair-mechanisms-for-maintaining-dna-344/#:~:text=DNA%20repair%20processes%20exist%20in,been%20highly%20conserved%20throughout%20evolution.
Posted by: guidrysaidence.blogspot.com
0 Response to "Does An Organism Have A Way Of Repairing Dna"
Post a Comment